



Дізнавайтесь першими найсвіжіші агрономічні новини України на нашій сторінці в Facebook, Telegram, а також підписуйтесь на Instagram СуперАгронома.
Раціональне використання поживних речовин для поліпшення властивостей кореневої системи і посухостійкості, — дослідження
5 травня 2021, 10:00

Коренева система сої
Наразі світова сільськогосподарська галузь переймається питанням: як досягти стійкої продовольчої безпеки за активізації кліматичних змін та почастішання посушливих сезонів. У зв'язку з цим дедалі більше уваги приділяють кореневій системі, оскільки саме коріння найбільш чутливо реагує на наявність поживних речовин і вологи. Припустивши, що за раціонального використання поживних речовин та води можна керувати властивостями кореневої системи культур задля підвищення посухостійкості, група вчених більш детально дослідила цей аспект та опублікувала результати своєї роботи на ресурсі researchgate.net.
Учені порівнювали вплив різних рівнів вологозабезпечення та поживних речовин на пластичність коріння та генетичне регулювання цих процесів, зосереджуючи увагу на тому, як це може вплинути на посухостійкість.
У довгостроковій перспективі науковці передбачають, що вплив на кореневу систему, зокрема на збільшення пластичності, сприятиме вирощуванню ефективних сортів, раціональне споживання ними вологи та поживних речовин — для досягнення стабільності у сільському господарстві.
Посухостійкість, холодостійкість, стресостійкість кукурудзи. Як запобігти втратам врожайності
ЧитатиПосухостійкість — це здатність рослин протистояти посусі, її часто розподіляють на такі складові: “порятунок” від посухи, її уникнення та толерантність. У своїй роботі науковці зосередились на здатності кореневої системи добувати воду навіть у пересушеному ґрунті, що є одним із аспектів уникнення посухи.
За час еволюції рослини навчились відчувати стан ґрунту, щоб регулювати споживання поживних речовин кореневою системою для підвищення більшої стійкості до посухи. Наразі є експериментальні докази та загальний консенсус щодо того, що кореневі сигнали регулюють реакції росту пагонів у відповідь на воду, появу поживних речовин та певні властивості ґрунту. Сигнали від кореня до пагона та від пагонів до кореня стають основним регулятором росту залежно від екологічних факторів.
МікроРНК, пептиди, абсцизова кислота (AБК), транс-зеатин, цитокінін, стриголактон та гібереліни є основними сигнальними молекулами, які регулюють ріст пагонів у відповідь на доступність води та поживних речовин, тоді як цитокінін, ауксин та цукор є ключовими сигнальними молекулами у зворотному напрямку — від пагона до коренів.
Кортикальні клітини в зоні подовження коренів відчувають рівень вологи у ґрунті, й під час посухи листя реагує закриттям продихів. Така реакція (тобто закриття продихів) може бути більш чутливою до рівня вологості біля коріння, і замість того, щоб втрачати значну кількість вологи через транспірацію, рослина закриває продихи задовго до зниження водного потенціалу листя, щоб уникнути кавітації ксилеми.
Цей зв’язок опосередковано здійснюється пептидом CLE25, який виробляється в коренях і рухається вгору до листя, де індукує синтез АБК, що спричиняє закриття продихів.
Брасиностероїди (BR) також синтезуються в судинній системі під час посухи та ініціюють накопичення осмопротекторів у коренях, які допомагають всмоктуванню води.
На архітектуру кореневої системи, тобто на розміщення коренів у тривимірному просторі, впливають як генетика, так і середовище.
Наприклад, інтрогресія (переміщення певного гена від одного виду до генофонду іншого через повторне зворотне схрещування міжвидового гібрида з одним із батьківських видів) гену DRO1, який відповідає за ріст кореня углиб, у рисову лінію з неглибоким корінням збільшила врожайність під час посухи завдяки подовженій кореневій системі, яка знаходила глибші запаси води.
Так само лінійка пшениці, що містить алель дикої тетраплоїдної пшениці, також продемонструвала глибше вкорінення та вищий урожай під час 4 посушливих періодів.

Однак в умовах достатньої вологи або періодичної посухи фенотиповий приклад росту коріння вглиб зі значною щільністю по довжині кореня може дорого обійтися, враховуючи витрати вуглецю на ріст та “утримання” тканин. Зокрема, система кореня, яка призначена для “дослідження” навколишнього ґрунту може споживати більш ніж 50% продуктів щоденного фотосинтезу. Є й інші побічні ефекти у різних фенотипів коренів. Наприклад, тривимірне (вшир) розгалуження підходить для розростання коренів у верхньому шарі ґрунту, але не придатне для видобування води з глибоких шарів, коли кількість води на нижніх рівнях значно збільшується під час дощового сезону.
З другого боку, надмірно розгалужена коренева система не завжди може бути придатна для забезпечення більш високого врожаю за низької вологості, адже значна кількість кореневих кінчиків може призвести до конкуренції між ними за вуглець, й вони не зможуть активно “досліджувати” ґрунт у пошуках вологи та поживи.
Для кращої посухостійкості за інтенсивних технологій пропонуються такі характеристики кореня:
- коротше осьове коріння,
- знижена бічна щільність коренів,
- знижена реакція на доступність ресурсів,
- більша повітряна паренхіма,
- зменшення кількості кортикальних клітин у більш крупних рослинах.
Враховуючи ці особливості та пластичність коренів, здатність генотипу змінювати фенотип в різних умовах може бути цінною перевагою і перспективним напрямом для селекції.
Слід також зазначити, що посуха також змінює мікробіоту в ризосфері. Водночас мікробні спільноти відіграють значну роль у відновленні рослини після посухи, а також стійкості рослин-господарів до стресів. Загалом же основна мета дослідження групи вчених — виявити можливості щодо підвищення стійкості до посухи шляхом управління пластичністю коренів за допомогою поживних речовин. Було обрано саме поживні речовини, оскільки внесення добрив — це широко застосовувана поліпшувальна стратегія проти посухи. Більше того, схожі механізми сприяють “захопленню” води та поживних речовин, отже, між сигнальними молекулами пошуку води та поживних речовин може бути перехресний обмін сигналами.
Що ж може відбуватися з корінням, залежно від умов у ґрунті?
Пластичність коренів у реакціях на вологу
Існує багато типів реакції коріння на дефіцит води. Наприклад, кореневий ксеротропізм — коли коріння у певній ділянці посилюють гравітропічну реакцію під впливом води, задля глибшого росту у пошуках вологи. Однак проникнення коренів на глибокі дистанції залежить як від властивостей ґрунту, так і від особливостей самих ренів. У сухій землі подовження коренів обмежене через механічні бар'єри та обмежений тиск тургору для подовження клітин кореня.
Як отримувати сталі врожаї кукурудзи в умовах посухи та дефіциту вологи
ЧитатиНеоднорідна доступність води у верхньому шарі ґрунту викликає викривлення коренів у напрямку до вологи, це називається гідротропізмом. Встановлено, що такі ознаки, як кількість кортикальних клітин, їх “старіння”, що пов'язано з утворенням кортикальної повітряної паренхіми (аеренхіми), забезпечують стійкість до посухи за рахунок росту кореня вглиб і при цьому низьких витратах вуглецю.
Наприклад, зниження осьової провідності коренів через малий діаметр ксилеми та менша кількість судин істотно збільшують потенціал урожайності пшениці в умовах посухи в Австралії.
Водночас у посухостійких генотипів рису спостерігаються великі судини ксилеми та менше утворення аеренхіми порівняно із вразливими щодо посухи генотипами. На відміну від рису, генотипи кукурудзи, що мають більшу кількість аеренхім, пов'язаних із посухостійкістю, досягають більшої компактності по довжині коренів і забезпечують вищу біомасу під час посухи.

Хоча багато з цих описаних ознак були вивчені в контексті генотипових відмінностей, деякі з них також виражаються, коли рослини перебувають у стресовому стані і тому стають пластичними при адаптації до посухостійкості.
Пластичність коренів у реакціях на поживні речовини
Рослини залежать від отримання багатьох поживних речовин із ґрунту, хоча ефективність сільського господарства здебільшого обмежується використанням азоту (N), фосфору (P) та калію (K). Таким чином, механізми коріеневих сенсорів та сигналів здебільшого досліджені на цих поживних речовина. Однак більша частина нашого розуміння сигналів поживних речовин походить від змодельованих рослин, тому сучасні знання рослинництва є обмеженими. У випадку використання азоту, зондування та відповіді можуть бути класифіковані як чотири механізми:
1) місцеве сприйняття та засвоєння поживних речовин у коренях;
2) сигнали коріння-пагін-коріння до частини кореня з низьким вмістом поживних речовин;
3) сигнали коріння-пагін-коріння до частини кореня з високим вмістом поживних речовин;
4) системний інгібуючий сигнал від пагона до кореня, щоб припинити живлення нутрієнтів, коли пагін вже має їх достатню кількість.
Амоній може стримувати подовження коренів, викликати їх бічне розгалуження або подовжувати кореневі волоски. Нітрат може чинити стримувальний або стимулювальний вплив на бічний ріст коренів, залежно від N-статусу рослини. За слабкого дефіциту підвищується бічний ріст коренів, тоді як за сильного бічний ріст коренів уповільнюється, а поява нових бічних коренів зупиняється. Нерівномірна доступність нітратів також стимулює довжину та щільність бічних коренів у насичених нітратами зонах.
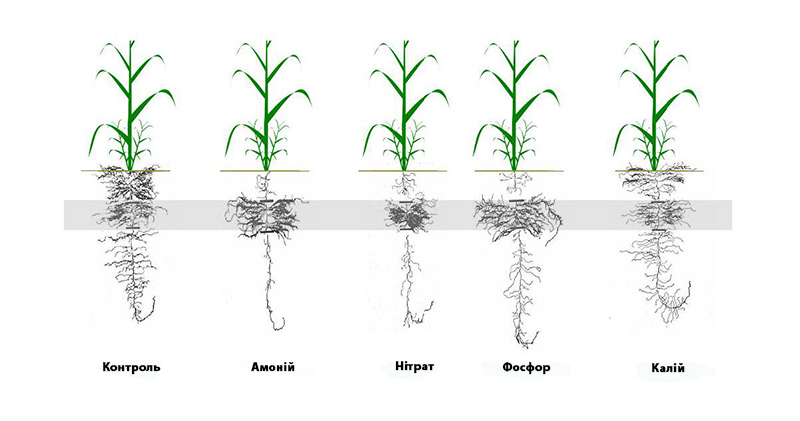
Рисунок 1: Архітектура кореневої системи ячменю під впливом локалізованого надходження поживних речовин (фосфат, 0,5 мг/л-1, нітрат 1,0 мг/л-1, амоній 1,0 мг/л-1 та калій 3,0 мг/л-1). На контролі рослини отримували 7 поживних речовин у всіх частинах кореня, тоді як при інших обробках лише там, де на малюнку затінені ділянки коренів (середня зона), отримували повноцінний поживний розчин, тоді як зверху та знизу забезпечувався дефіцит зазначених елементів.
Загалом, бічні корені більш чутливі до зміни кількості поживних речовин, ніж осьові. Більш товсте коріння менш схильне до зневоднення, має вищу гідропровідність, але у багатьох культурах спричиняє більший вихід вуглецю і має нижчі пластичні властивості в реакціях на доступність поживних речовин. У просторово гомогенному стані з низьким вмістом азоту стимулюється бічний ріст, тоді як при гомогенному високому стані бічний ріст обмежується. Однак, коли в ґрунті з низьким вмістом азоту з'являється ділянка з високим вмістом азоту, бічні корені здатні розростатися й розгалужуватися в цій невеликій ділянці. Вища доступність фосфатів збільшувала довжину первинного кореня з більш рідкими та укороченими бічними корінцями.
У ріпаку, томатів та шпинату низький рівень фосфору спричиняє утворення довгих кореневих волосків з високою щільністю убік напрямку, де вищий рівень фосфору.
Пластичність коренів у взаємопов'язаних реакціях на наявність вологи та поживних речовин
Незважаючи на те, що застосування добрив є загальноприйнятою агрономічною практикою для посилення росту рослин та стійкості до посухи, наразі дуже мало уваги приділяється тому, як місцеве та своєчасне внесення поживних речовин може вплинути на просторовий та часовий розподіл коріння для подальшого підвищення посухостійкості. Як правило, реакції пластичності коренів вивчали на дефіцит води або 8 поживних речовин без взаємодії, і часто в контексті інтенсивного землеробства. Звісно, ці дослідження поліпшують розуміння того, як рослини регулюють кореневу архітектуру у просторі та часі, але вони не мають критичного розуміння того, наскільки багато поживних речовин можна використовувати для її оптимізації для посухостійкості.
Догляд за посівами озимих зернових восени: акцент на розвиток кореневої системи
ЧитатиНаприклад, в одному з польових експериментів стан посівів пшениці, які потерпали через зменшення доступності вологи, було поліпшено за допомогою локального внесення добрив (NPK) на глибині 8-10 см. Порівняно з посівами, де розподіл добриви вносились суцільно, пшениця з локальним внесенням мала вищий показник біомаси — завдяки тому, що споживання води та інших поживних речовин здійснювалось навіть тим корінням, що розміщувалось поза зоною, насиченою поживними речовинами, і сягало значно глибше.
В одному з досліджень було виявлено, що на обсяг та щільність коріння впливає кальцій та взаємодія з ним вологи. А саме — за вищих рівнів K зменшення площі коренів та їх щільності були мінімальними порівняно з низьким вмістом K за тих же рівнів вологи. Встановлено, що вищий вміст кальцію поліпшує максимальну ширину кореневої системи та глибину вкорінення під впливом вологи. Водночас результати свідчать і про те, що за значного дефіциту вологи вплив від рівня калію був не таким вагомим.

Підвищення стійкості до посухи за допомогою доцільного застосування поживних речовин
За інтенсивних технологій управління живленням спрямоване на забезпечення достатньої та рівномірно розподіленої кількості елементів поблизу поверхні ґрунту, де коріння виходять із насіння або стебла. Однак, враховуючи той факт, що коріння здебільшого розростається в ділянках із високим вмістом поживних речовин, ця стратегія насправді може перешкоджати здатності рослини пускати глибші корені, що в подальшому могло б забезпечити кращу посухостійкість. Водночас рослини з достатнім мінеральним живленням витрачають менше нутрієнтів на кореневу систему, ніж на верхню частину, що теж може перешкоджати посухостійкості.
Деякі підходи до оптимізації застосування добрив
ЧитатиНатомість розміщення на глибині та роздільне внесення добрив може бути використано для стимулювання росту коріння углиб.
Глибоке розміщення добрив вже застосовується для підвищення ефективності використання поживних речовин і може бути додатково оптимізовано для формування кореневої системи, яка краще поглинатиме вологу.
У кукурудзи більша щільність по довжині кореня за розростання тонких корінців була досягнута завдяки застосуванню локалізованого розміщення аміаку та фосфору, що призводило до збільшення зеленої маси та споживання поживних речовин. Також відомо, що такий спосіб збільшення кореневої системи може поліпшити посухостійкість кукурудзи.

Отже, таке розміщення та оптимізація поживних речовин можуть бути корисними для підвищення стійкості сільськогосподарських культур до посухи (рис. 2).
Загалом реакція коренів на локалізоване розміщення поживних речовин відрізнялась між культурами та генотипами через різний рівень пластичності коренів. Зокрема, злаки проявляють більшу пластичність коренів, ніж бобові. Більше того, норму удобрення поля слід врегульовувати з урахуванням наявності вологи протягом вегетації, оскільки дифузія іонів від глибокого розміщення добрив може травмувати верхні меристеми коренів під час посухи та негативно вплинути на надземну частину.
Оскільки надмірне застосування поживних речовин знизить їх оптимальний розподіл і доступ до кореневої системи, вчені також припустили, що роздільне внесення елементів живлення може не тільки підвищити ефективність їх використання, а й сприяти глибшому росту й розвитку коріння і як наслідок — стійкості до посухи.
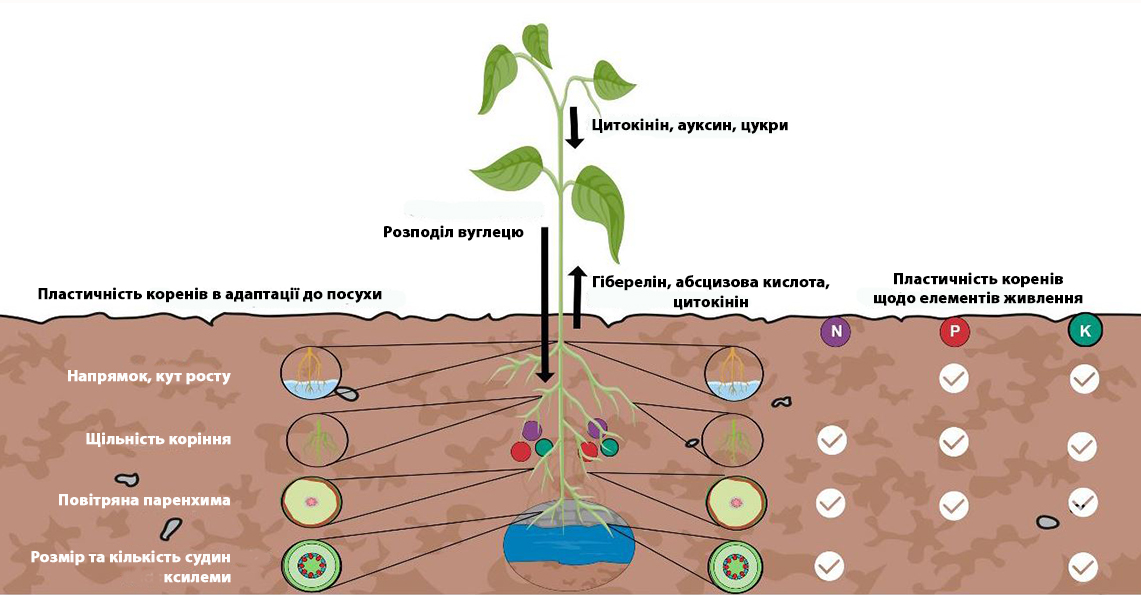
Рисунок 2: Раціональне просторове та часове розміщення поживних речовин може допомогти в оптимізації властивостей кореневої системи, пристосованих до посухи, для збільшення доступу до вологи і провідності води, що збільшує загальну життєздатність рослин.
Майбутні перспективи
Різні типи кореневих систем по-різному реагують на воду та поживні речовини.
Однак розуміння взаємодії вологи та поживних речовин при формуванні кореневої системи буде надзвичайно важливим для оптимізації норми добрив відповідно до наявності вологи.
Учені запропонували чотири стратегії для раціонального застосування поживних речовин, що може оптимізувати кореневі властивості при дефіциті води:
1) вплив на підвищення загальної енергії та ріст рослин, зокрема цілеспрямована дія на ріст кореневої системи,
2) збільшення розподілу вуглецю в кореневій системі за рахунок зменшення співвідношення пагона до кореня,
3) вплив на конкретні властивості коріння, зокрема, напрямки росту, кількість відгалужень, діаметри та довжина коренів, властивості ксилеми, — завдяки використанню відповідних норм та співвідношень поживних речовин,
4) застосування технології глибокого розміщення добрив для стимулювання розростання коренів на глибині.
Подальші дослідження варто спрямувати на вивчення регуляторних молекулярних механізмів, що впливають на реакції рослин, та напрацювання ефективних агрономічних практик і розробку технічних засобів для економічного, раціонального внесення добрив.
Читайте також

Нові агрегати для обробітку проміжних культур та післяжнивних решток виводить на ринок України чеський виробник

Дощі по всій території України поповнили вологозапаси в ґрунті до достатніх та оптимальних показників

Вперше в історії колообіг води на планеті порушено: це одна з причин посух та втрат у рослинництві

Накопичена ґрунтова волога врятувала врожайність 2024, наступного року без неї будуть проблеми — Сергій Авраменко
Матеріали до теми



Популярне


